|
If enough oxygen is present, then pyruvate diffuses through the
two membrane layers into the mitochondrial matrix and enters the
cit ric acid cycle. The enzymes of this cycle all are dissolved
in the matrix except for three: succinate, pyruvate, and a-ketoglutarate
dehydro genase. Succinate dehydrogenase is the only enzyme that
transfers hydrogen atoms to FAD rather than NAD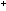 .
The succinate dehydrogenase molecule must be embedded in the inner
mitochondrial membrane alongside the respiratory cytochromes and
enzymes, because its carrier molecule, FADH .
The succinate dehydrogenase molecule must be embedded in the inner
mitochondrial membrane alongside the respiratory cytochromes and
enzymes, because its carrier molecule, FADH ,
is permanently bound to the enzyme and cannot diffuse from one place
to another as can NADH. Pyruvate and ,
is permanently bound to the enzyme and cannot diffuse from one place
to another as can NADH. Pyruvate and 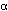 -ketoglutarate
dehydrogenase both are large, multienzyme complexes with molecular
weights in the millions, and are similarly embedded in the inner
mito chondrial membrane. The other citric acid cycle enzymes float
freely in the matrix. NADH produced in the cycle diffuses to the
inner mem brane surface, where it and FADH.
are reoxidized by the respiratory chain. Oxygen is reduced to -ketoglutarate
dehydrogenase both are large, multienzyme complexes with molecular
weights in the millions, and are similarly embedded in the inner
mito chondrial membrane. The other citric acid cycle enzymes float
freely in the matrix. NADH produced in the cycle diffuses to the
inner mem brane surface, where it and FADH.
are reoxidized by the respiratory chain. Oxygen is reduced to 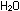 ,
and ADP is phosphorylated to ATP, both processes occurring at the
inner membrane surface. ,
and ADP is phosphorylated to ATP, both processes occurring at the
inner membrane surface.
|
|
It is the inner membrane that isolates the mitochondrion chemically
from the cell in which it sits. The outer membrane is permeable
to most molecules of low molecular weight. The inner membrane will
allow only water, small neutral molecules, and short-chain fatty
acids to pass through. It is impermeable to cations and anions,
most amino acids, sucrose and other sugars, coenzyme A and its esters
with acetate and succinate, and ADP, ATP, NAD,
and NADH. Some of these molecules are transported back and forth
by carrier molecules, or permeases.
One such permease exchanges ADP and ATP across the inner membrane
on a one-for-one basis. Other shuttle molecules can transport fatty
acid-coenzyme A complexes, phosphate, hydroxide ion, citrate, isocitrate,
succinate, and malate, but not oxaloacetate. This impermeability
of the inner membrane to oxaloacetate is the reason for the conversion
of oxaloacetate to malate and back again during gluconeogenesis.
|